Biologie
Nutrition – Reproduction – Habitat – Distribution géographique – Cartes : Progression – Prédictions
Activité, comportement
Vespa velutina est une espèce diurne qui, contrairement au Frelon d’Europe, V. crabro, interrompt toute activité à la tombée de la nuit. C’est un prédateur avéré d’autres Hyménoptères sociaux, notamment des abeilles et des guêpes communes mais, comme V. crabro, il consomme aussi une grande variété d’autres insectes et d’araignées (Villemant et al., 2006b ; Perrard et al., 2009 ; Rome et al., 2011 ; Villemant et al., 2011b ; Muller et al., 2013).
Depuis l’été 2006, certains apiculteurs s’inquiètent de voir leurs ruches plus régulièrement et plus fortement attaquées par cette espèce que par le Frelon européen. En vol stationnaire à une vingtaine de centimètres de l’entrée de la ruche, une ouvrière de V. velutina succède régulièrement à une autre pour capturer les butineuses qui reviennent chargées de pollen. Le frelon fonce sur sa proie, la saisit entre ses pattes et la tue d’un coup de mandibules derrière la tête avant de l’emporter dans un arbre pour la dépecer. Il ne conserve que le thorax contenant les muscles du vol riches en protéines ; il en fait une boulette qu’il emporte jusqu’au nid pour en nourrir les larves (Janet, 1903 ; Rome et al., 2015).

En Asie, Vespa velutina est considéré comme un redoutable ennemi des ruchers (Shah & Shah, 1991 ; Ken et al., 2005) car il peut détruire jusqu’à 30 % d’une colonie de l’Abeille asiatique, Apis cerana. Après avoir décimé les gardiennes, les ouvrières du Frelon peuvent pénétrer dans la ruche pour prélever le couvain (Arca et al., 2014). En France, la conformation des ruches permet de réduire l’entrée à une étroite fente, ce qui interdit la pénétration des insectes d’une taille supérieure à celle des abeilles. La prédation exercée par V. velutina se limite alors aux abeilles adultes, mais sa présence insistante, parfois en grand nombre (15 à 20), devant les ruches stresse les abeilles, réduisant leur sorties, ce qui limite les récoltes de nectar et de pollen à un moment où les abeilles élèvent leurs dernières ouvrières de l’année et font leur réserve pour passer l’hiver (Requier et al., 2019).

En France, tous les observateurs s’accordent sur le fait que V. velutina n’est pas agressif et qu’il est possible d’observer son nid à 4 ou 5 m de distance sans risque. Les rares personnes piquées l’ont été en tentant de détruire un nid ou en touchant une ouvrière par inadvertance. La piqûre, si elle est douloureuse, n’est pas plus dangereuse que celle d’une guêpe ou d’une abeille mais les personnes allergiques au venin d’Hyménoptères doivent bien sûr rester très prudentes (De Haro & Blanc-Brisset, 2009 ; De Haro et al., 2010 ; Schwartz et al., 2012 ; Viriot et al., 2015).
Plusieurs personnes ont côtoyé des nids en activité installés sur ou à proximité de leur habitation sans que les ouvrières ne manifestent une quelconque agressivité lors des allées et venues des habitants (Villemant et al., 2006b). Il faut toutefois demeurer extrêmement prudent face aux très gros nids installés dans les arbres. Lorsque l’on s’approche à moins de 5 m d’un nid de Frelon, plus la colonie qu’il renferme est importante et plus les risques sont grands de subir l’attaque d’un essaim d’ouvrières.
Comportement – Reproduction – Habitat – Distribution géographique – Cartes : Progression – Prédictions
Nutrition
Comme les autres frelons, Vespa velutina est un prédateur généraliste qui s’attaque à une très grande variété de proies. Outre les abeilles, il s’attaque aux insectes de très nombreux ordres et notamment aux guêpes, mouches, papillons ainsi qu’aux araignées (Muller et al., 2009 ; Rome et al., 2011b ; Muller et al., 2013). Il prélève aussi de la viande sur des cadavres de vertébrés ou sur les poissons et crevettes exposés aux étals des marchés. Les boulettes de proies servent à nourrir les larves. Les adultes ne se nourrissent que de liquides sucrés (miellat, nectar, miel…) et du liquide riche en protéines que régurgitent les larves lorsqu’ils les sollicitent (Janet, 1903 ; Matsuura & Yamane, 1990). Les ouvrières transportent ces liquides dans leur jabot pour nourrir par trophallaxie la reine, les autres ouvrières, les mâles et les futures fondatrices restées dans le nid. À l’automne, ils mangent aussi la chair des fruits mûrs, pommes, prunes, raisins…

Comportement – Nutrition – Habitat – Distribution géographique – Cartes : Progression – Prédictions
Reproduction
C’est en automne (octobre à novembre) que les femelles reproductrices de la nouvelle génération quittent le nid en compagnie des mâles pour s’accoupler ; elles sont les seules à hiverner tandis que les mâles, les dernières larves et les ouvrières meurent. Au printemps (mars à juin), chaque reine fondatrice ébauche un nouveau nid, pond quelques œufs et soigne ses premières larves qui deviendront, un mois à un mois et demi plus tard – selon la température -, des ouvrières adultes capables de prendre en charge la construction du nid et l’entretien de la colonie. La reine consacrera alors le reste de sa vie à pondre. Avec l’apparition des ouvrières, l’activité de la colonie s’intensifie considérablement et la taille du nid augmente pour atteindre son maximum au début de l’automne (Rome et al., 2013b ; Rome et al., 2015, Villemant & Rome, 2017).

Comme chez tous les autres Hyménoptères, les descendants femelles sont issus d’œufs fécondés et les mâles d’œufs non fécondés. La colonie n’est composée que d’ouvrières (femelles stériles) jusqu’à ce que la nouvelle génération de sexués mâles et femelles se développe au début de l’automne. La vieille reine meurt peu de temps avant l’essaimage des sexués. Le reste de la colonie dépérit et meurt au cours de l’hiver. Quelques rares nids peuvent demeurer actifs en décembre. Les nids vides ne sont jamais réutilisés mais on y trouve parfois au début du printemps quelques femelles sexuées tardives qui sont restées bloquées par l’arrivée du froid. Elles sont incapables de fonder une colonie car elles n’ont pas été fécondées et ont souvent des ailes atrophiées.
Les nids apparaissent en moyenne cinq fois plus populeux que ceux du Frelon d’Europe. Les plus grands peuvent produire plus de 13 000 individus au cours de la saison (d’avril à novembre) et peuvent contenir à l’automne près de 2 000 ouvrières qui élèvent au moins 500 futures fondatrices, mais probablement plus d’un millier, et autant de mâles (Rome et al., 2015).
Les femelles fécondées hivernent isolément ou par groupes de deux ou trois dans la litière ou les troncs pourris ; certaines reprennent leur activité dès le mois de février. Les jeunes nids, de la taille d’une orange, sont installés à partir du mois de mars, sur le rebord d’un toit, dans divers abris ou des ruchettes vides. Ils comptent une dizaine de cellules entourées d’une fine coupole de papier puis d’une enveloppe sphérique (Villemant & Rome, 2017). Les premières ouvrières, qui ne mesurent qu’entre 14 et 16 mm, émergent au cours du mois de mai. 70 % des colonies déménagent, dans le courant du mois d’août, lorsque le nid primaire est placé trop près du sol ou dans un endroit confiné ; la colonie s’installe alors dans un nouveau nid construit par les ouvrières souvent à plus de 10 m dans un arbre (Rome et al., 2015).
Comportement – Nutrition – Reproduction – Distribution géographique – Cartes : Progression – Prédictions
Habitat
Dans son aire d’origine, le Frelon asiatique vit essentiellement en milieu forestier mais avec l’extension de l’urbanisation aux dépens de la forêt, on le rencontre aussi en zone périurbaine (Yamane, 1977 ; Nakamura & Sonthichai, 2004 ; Sheng & Xing, 2005). En climat subtropical, la variété nigrithorax se cantonne généralement aux régions montagneuses (Van der Vecht, 1957).
En France, le Frelon asiatique installe de préférence son nid dans les hautes branches des grands arbres, en zone urbaine ou agricole comme en milieu boisé (Rome et al., 2009 ; Rome et al., 2015). Il semble profiter des vallées des cours d’eau et des grands axes routiers pour se disperser ; il semble par contre éviter les peuplements purs de conifères. En effet, bien qu’il soit présent dans le département des Landes, les forestiers n’ont que rarement repéré de nid en pleine forêt.

Parmi les 14794 nids correctement géo-référencés entre 2004 et 2016, près de 53% étaient localisés dans une zone urbaine ou péri-urbaine, 38% en milieu agricole, plus 8% en forêt et zones naturelles (prairies, landes…) et moins de 1% en milieu humide (Villemant et al., 2011a ; Rome et al., 2015 ; Rome, 2015 ; Fournier et al. 2017).
Comportement – Nutrition – Reproduction – Habitat – Cartes : Progression – Prédictions
Distribution géographique
Vespa velutina a été décrite par Lepeletier en 1836 à partir de spécimens collectés dans l’île de Java (Indonésie). Sa coloration est extrêmement variable : on distingue une douzaine de variétés, dont la variété nigrithorax qui a été décrite de Darjeeling (Inde) en 1905 par du Buysson (Carpenter & Kojima, 1997 ; Perrard et al., 2014).

L’aire d’origine de l’espèce Vespa velutina s’étend de l’Afghanistan à la moitié sud de la Chine, Taiwan compris, et de la péninsule indochinoise (Thaïlande, Laos, Vietnam) à la Malaisie et à l’archipel indonésien. La variété V. velutina nigrithorax vit au nord de l’Inde (Darjeeling, Sikkim), à l’est du Népal, au Bhoutan et en Chine (Carpenter & Kojima, 1997 ; Rome et al., 2010 ; Rome et al., 2011b ; Villemant et al., 2011a). Elle a été signalée pour la première fois en Corée en 2006 (Kim et al., 2006 ; Choi et al., 2012). En Asie continentale, la variété nigrithorax vit sous des climats comparables à ceux de l’Europe.
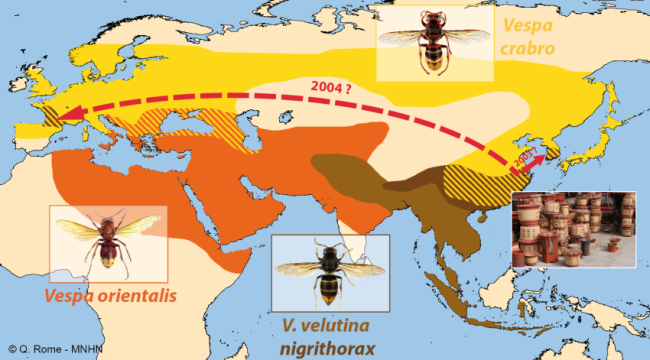
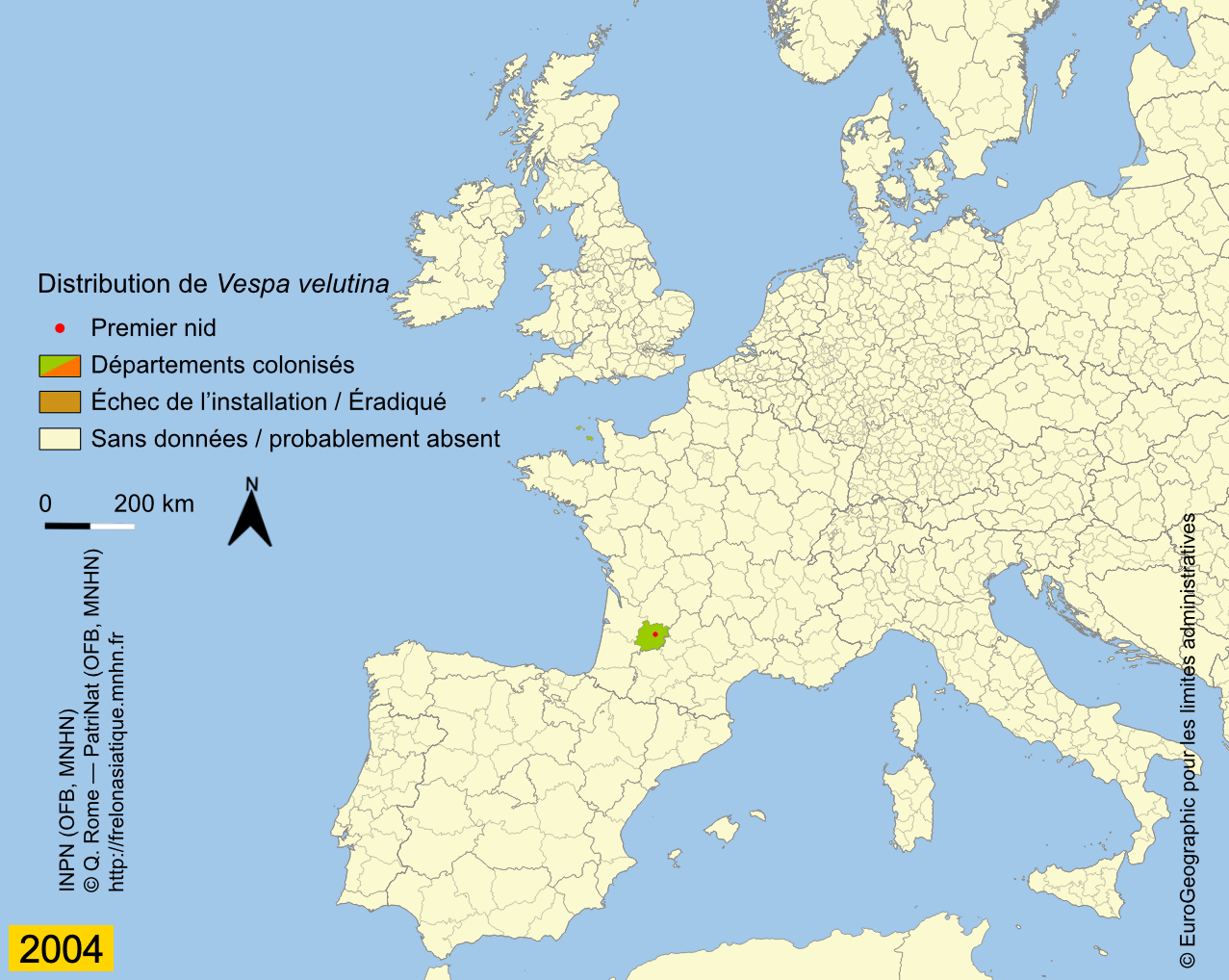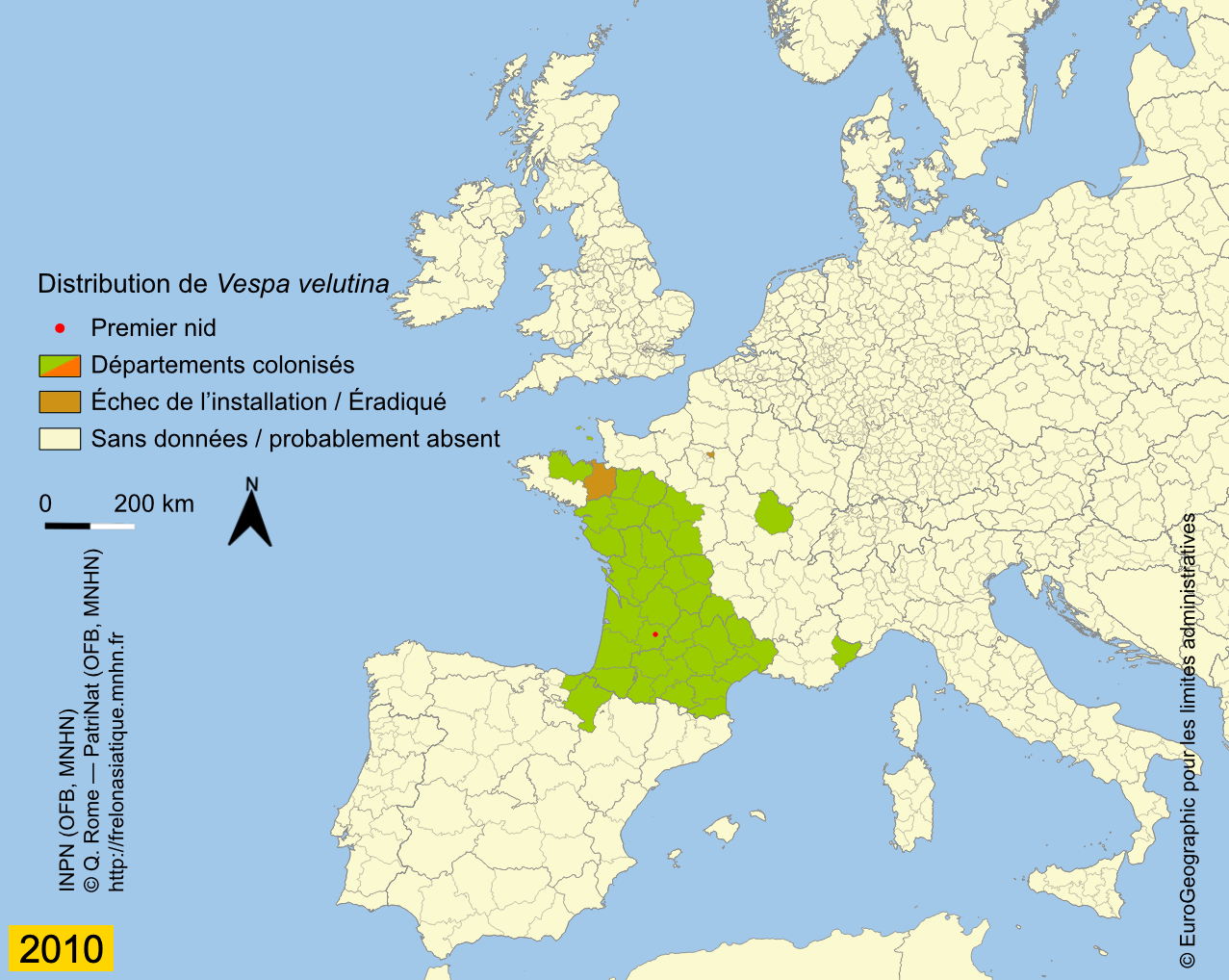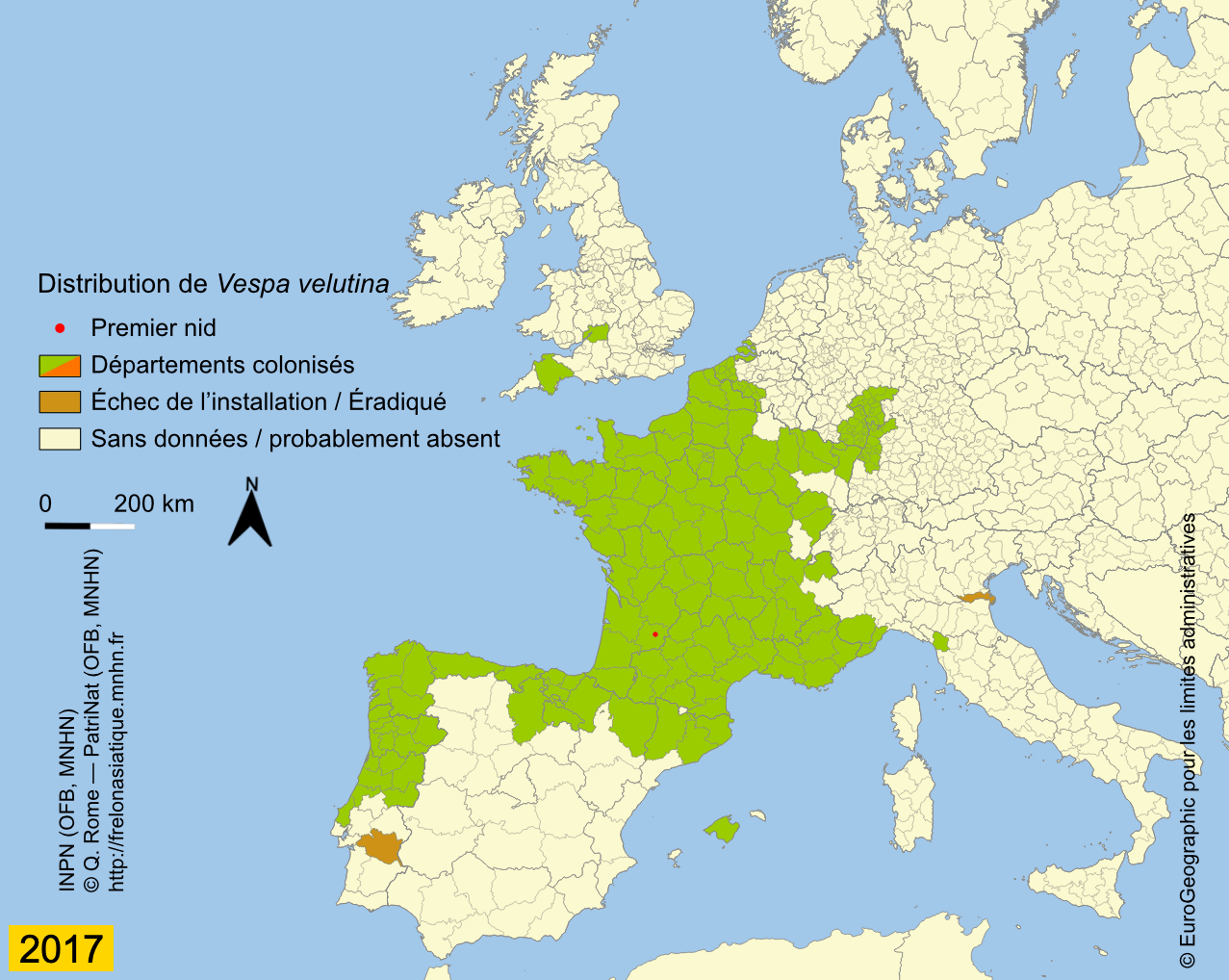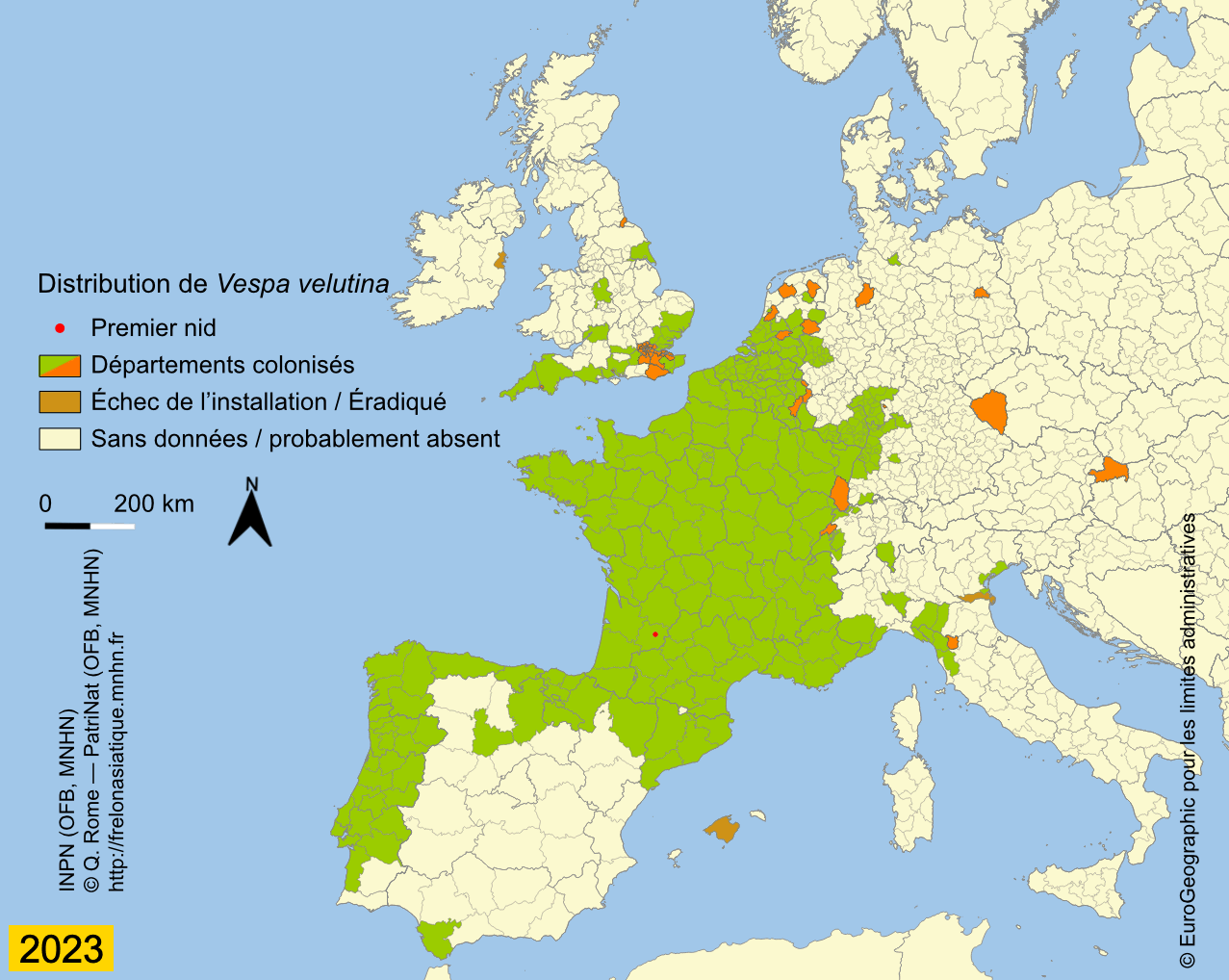
Depuis son introduction en France, probablement un peu avant 2004, l’expansion de Vespa velutina a été très rapide. La carte des départements envahis, régulièrement mise à jour sur le site de l’INPN, montre que le front d’invasion progresse en moyenne de 78 km par an (Rome et al., 2015 ; Robinet et al., 2017). Les deux premiers nids ont été trouvés dans le Lot-et-Garonne en 2004 et déclarés, après destruction au fusil, à la SRPV de Tonneins. Des individus ont été par la suite capturés durant l’hiver 2005 ainsi qu’un nid observé. Des captures de fondatrices au printemps 2006 et le lancement de la surveillance ont confirmé l’installation de l’espèce, déjà présente dans 13 départements (Haxaire et al., 2006 ; Villemant et al., 2006b). Le nombre de nids observés s’est ensuite accru progressivement pour atteindre 1637 nids dans 32 départements en 2009 (Rome et al., 2009). Depuis l’enregistrement des nids se fait en priorité dans les nouveaux départements envahis car cela demande un important travail de vérification à partir des photos demandées aux observateurs. En effet, un tiers environ des signalements s’avère erroné du fait des confusions avec les nids du Frelon d’Europe ou d’autres espèces de guêpes (voire avec des nids de Pie) ou parce qu’ils correspondent à des doublons (un même nid signalé par plusieurs observateurs). Prendre en compte ces signalements erronés conduirait à une surestimation importante de l’extension actuelle de l’invasion (Rome et al., 2011b). Le suivi de l’invasion montre cependant que 56 départements étaient déjà envahis en 2012 sur une surface d’environ 345 000 km² et qu’en 2014, le frelon était présent dans 67 départements, soit environ 360 000 km². L’invasion dépasse maintenant nos frontières puisque le frelon se répand au nord de l’Espagne (Pays basque, Navarre) depuis 2010 (López et al., 2011) ; il a été aussi introduit accidentellement en 2011 au Portugal (Grosso-Silva & Maia, 2012), en Galice et en Catalogne (Espagne) (Goldarazena et al., 2015). Observé aussi en 2011 dans le département français du Nord et en Belgique, le frelon n’a pas été revu depuis, ce qui laisse penser qu’il n’a pas réussi pour l’instant à s’y acclimater (Rome et al., 2013a). En 2012 il s’implante en Italie (Porporato et al., 2014), en 2014 en Allemagne (Witt, 2015), au Royaume-Uni et en Belgique en 2016 (Keeling et al., 2017 ; Garigliany et al., 2017), enfin aux Pays-Bas en 2017 (Waarneming, 2017). Vespa velutina a également été introduit avant 2003 en Corée du Sud (Kim et al., 2006) et au Japon en 2012 (Ueno, 2014).
Comportement – Nutrition – Reproduction – Habitat – Distribution géographique – Carte : Prédictions
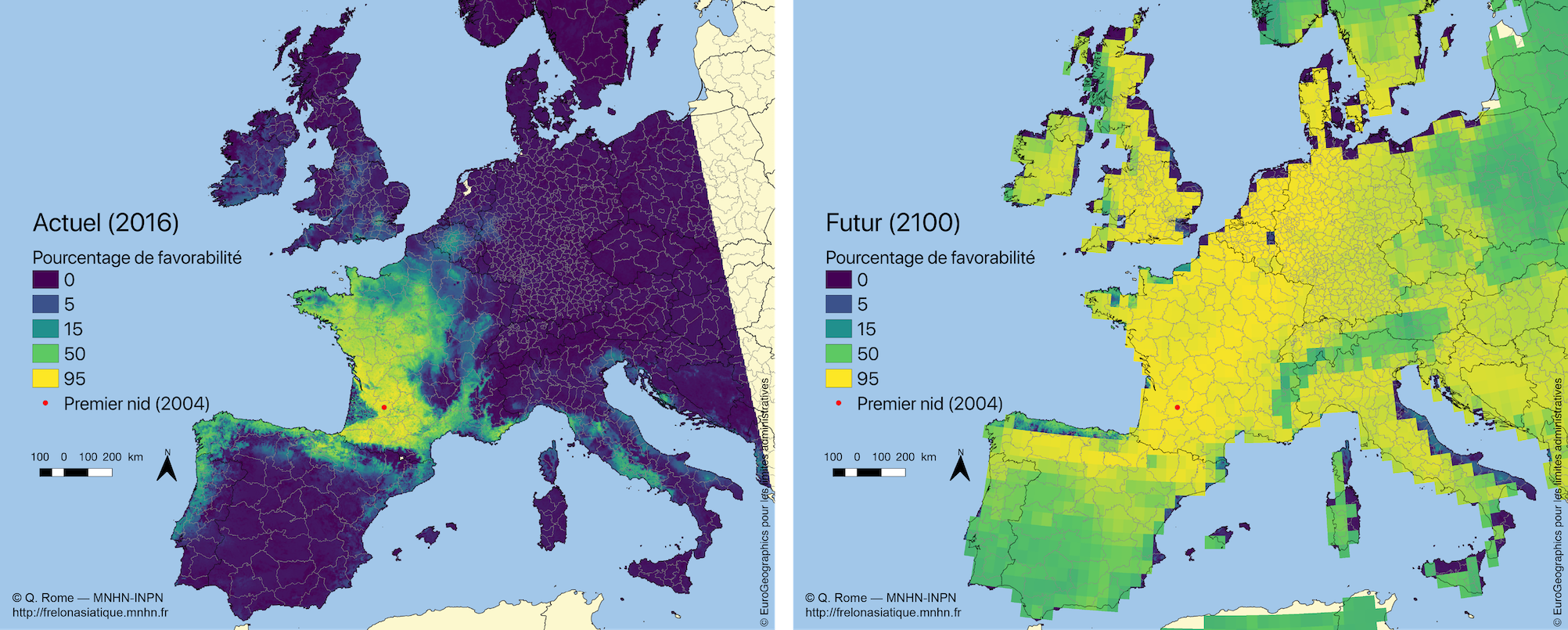
L’extension actuelle de l’invasion, consultables avec toute la traçabilité sur l’INPN, conforte les résultats des modélisations (niches climatiques) effectuées en 2010 à partir des données de présence de la variété envahissante, V. velutina nigrithorax, dans son aire d’origine comme dans l’aire d’invasion (France et Corée) (Barbet-Massin et al., 2018).
Ces modélisations montrent que la plupart des pays d’Europe ont un risque non négligeable de voir le frelon s’acclimater. Il en est de même de nombreuses autres régions du monde où l’espèce pourrait se maintenir en cas d’introduction accidentelle. À l’échelle du monde, la distribution potentielle du Frelon asiatique se rapproche de celle de la Guêpe européenne Vespula germanica. Cette guêpe envahissante introduite via le commerce international a en effet colonisé de vastes territoires dans l’hémisphère sud, où elle provoque localement de graves perturbations de l’environnement (Villemant et al., 2011a). Les récents scénarios de réchauffement climatique font craindre une expansion future encore plus rapide que celle des dix dernières années (Barbet-Massin et al., 2013 ; Rome et al., 2014).
Comportement – Nutrition – Reproduction – Habitat – Distribution géographique – Carte : Progression